






Per comprendere l'organizzazione interna dell'encefalo e lo sviluppo del sistema visivo, occorre considerare la sua origine embrionale.
La formazione del Sistema Nervoso avviene già dall'impianto dell'embrione nella parete uterina, quando le cellule iniziano a migrare e formare uno strato esterno di cellule ecrodermiche. Successivamente, nel corso della 2°-3° settimana post-concezionale inizia a differenziarsi la cosiddetta placca neurale: dalla superficie ectodermica dorsale si sviluppa una concavità sulla linea mediana che si unisce in prossimità dei bordi per formare il tubo neurale, contentente un fluido all'interno della sua cavità, noto come neurocele.
In queste prime fasi iniziano i primi processi di differenziazione dei neuroni, che proseguirà fino all'età adulta.
Nelle settimane seguenti l'estremità craniale del tubo comincia a modificarsi, dando forma a tre espansioni, dette vescicole encefaliche primarie, che in base alla loro posizione vengono denominate proencefalo, mesencefalo e romboencefalo. Attraverso successive trasformazioni, queste porzioni daranno origine agli emisferi cerebrali e al diencefalo, al mesencefalo, al ponte, al cervelletto e posteriormente permetteranno la formazione del midollo spinale.
Questo processo, di neurulazione, si completa entro la 6° settimana, a cui seguirà un priodo di proliferazione e migrazione cellulare con risultato finale la struttura completa del cervello, tronco cerebrale e midollo spinale.
Le funzioni visive, e sensoriali, nel bambino si modificano rapidamente nel corso dei primi mesi di vita, e raggiungono una stabilità nel periodo che va tra i 6 mesi fino ai 10-12 anni di età.
I danni che si verificano all'apparato visivo nei primi sei mesi di età possono comportare dei danni irreversibili; dopo i sei mesi, possono invece generare un regresso di quelle facoltà visive acquisite fino a quel momento, che possono in un certo qual modo essere recuperate grazie ad un intervento riabilitativo precoce.
E' nei primi mesi di vita infatti, che iniziano a svilupparsi le principali funzioni monoculari e binoculari, sia a livello sensoriale che motorio, la convergenza, l’accomodazione e i movimenti saccadici orizzontali.
Nella tabella sottostante vengono schematizzate le funzioni visive emergenti nelle diverse fascie di età, dalla 0 a 48 mesi.
|
0-1 mese |
|
||
|
1-2 mesi |
|
||
|
2-3 mesi |
|
||
|
3-4 mesi |
|
||
|
4-5 mesi |
|
||
|
5-6 mesi |
|
||
|
6-7 mesi |
|
||
|
7-8 mesi |
|
||
|
9-10 mesi |
|
||
|
11-18 mesi |
|
||
|
18- 24 mesi |
|
||
|
24-30 mesi |
|
||
| 30- 36 mesi |
|
||
| 36-48 mesi |
|
||
Tabella 1. Sviluppo delle funzioni visive nelle diverse fasce di età.
Quando si parla di acuità visiva, si è soliti intendere la misura del più piccolo dettaglio che può esssere individuato dal sistema visivo. Fisiologicamente, in un bambino di tre mesi viene stimata intorno ai 3 cicli/grado, e migliora nel corso dei primi mesi grazie ai processi di mielinizzazione delle vie ottiche ( più rapidi nei primi tre mesi di vita), l'aumento della proliferazione cellulare e del numero delle sinapsi nei primi sei mesi, e la formazione della fovea con successivo incremento dei coni al suo interno.
La stessa linee evolutiva la si risconta nello sviluppo della sensibilità ai contrasti, cioè l'abilità di differenziare la luminosità fra parti diverse di un'immagine.
Molti studi ( cfr Slater and Sykes, 1977 ) attestano che nel bambino alla fine del primo mese di vita è presente la capacità di discriminare l'orientamento di linee nello spazio (orizzontali e verticali) e i colori.
Successivamente, come testimoniato dallo studioso Fantz (1961-1963), è stato dimostrato che un bambino intorno ai due mesi è in grado di discriminare il volto umano da uno stesso volto deformato, cosa non riscontrabile nel bambino di un mese: ciò è dovuto ai circuiti cerebrali che diventano fuzionali fra il primo e il secondo mese di vita.
Per quanto riguarda la visione dei colori, a circa 8 settimane un bambino distingue meglio sfondi con grosse forme di colori contrastanti; verso le 16 settimane riesce a cogliere anche forme e righe piu sottili; verso le 20 settimane inizia a distinguere diversi colori più vivaci; infine, intorno alle 30 settimane, la visione si completa con i dettagli.
Invece, le strutture sottocorticali appaiono mature molto precocemente: queste sottendono l' abilità di percepire il movimento di un oggetto, non tanto la direzione, funzione specifica delle cellule corticali striate ed extrastriate, che compare a partire dal terzo mese.
Inoltre, grazie allo studio dei Potenziali Visivi Binoculari, si è concordi nel ritenere che intorno al terzo-quarto mese venga raggiunta la visione binoculare.
Il campo visivo di un neonato è notevolmente ridotto lateralmente, ed è con il trascorrere dei primi mesi che il range aumenta, fino al sesto mese, quando il bambino è in grado di spostare il campo visivo ( o meglio l 'attenzione visiva) anche lateralmente in uno spazio più ampio, come un adulto. Precisamente, all'inizio del secondo mese di vita, un bambino esplora la figura in modo frammentato, soffermandosi solo sui contorni esterni. E' dal terzo mese di vita che il bambino organizza i pattern di scansione oculare ed inizia a discriminare anche le parti interne di una figura, come i diversi elementi di un volto umano, grazie al funzionamento dei circuiti cerebrali, come sopra citato. (Salapatek, 1975)
Per quanto riguarda i movimenti rapidi, i saccadici, eseguiti per portare l'immagine dell'oggetto sulla fovea, sono presenti fin dalla nascita, anche se si tratta di movimenti di ampiezza standard e ipometrici, non adeguati alla distanza del target da fissare, come avviene nell'adulto: ciòè è dovuto alla mancanza dei meccanismi di programmazione dei saccadici che maturano nei mesi successivi al primo.
Grazie ai movimenti saccadici, il neonato di un mese è in grado di seguire il movimento di un oggetto, ma non di mantenere la fissazione continua sull'oggetto in movimento, che compare
a partire dal secondo mese. Secondo alcuni studi questo potrebbe essere dovuto a un deficit attentivo più che all'immaturità del sistema oculomotore. ( Atkinson, 1984)
I primi sei mesi di vita appaiono così relativi per quanto riguarda lo sviluppo e la maturaione delle funzioni visive, che raggiungono i livelli dell'adulto e faavoriscono la stabilità delle future competenze in ambito cognitivo, percettivo ed integrazione sociale.
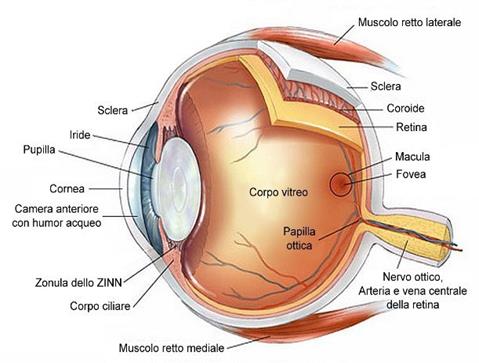
Fig.1 - Anatomia dell'occhio
L'occhio è l'organo deputato alla visione e viene considerato l'interfaccia del sistema nervoso sull' ambiente esterno. Esso è un esterocettore, in quanto capta gli sstimoli provenienti da fuori l'organism; in particolar modo si tratta di un telecettore, perchè raccoglie forme di energia che vengono da lontano.
Ogni occhio ha la forma di una sfera leggermente irregolare, con un diametro medio di circa 24mm e pesa circa 8g.
Le sue strutture principali sono: la cavità orbitaria, il bulbo oculare e le vie ottiche.
La cavità orbitaria è una cavità ossea che avvolge e contiene l'occhio ed i suoi annessi. Nella sua porzione anteriore si trovano il bulbo oculare, le palpebre e le vie lacrimali; la porzione posteriore contiene del tessuto adiposo che avvolge i muscoli, i vasi, i nervi propri dell'occhio.
Il bulbo oculare, di forma grossolanamente sferica, risulta composto da tre strati, tre tuniche che, andando dall'esterno verso l'interno, sono:
Inoltre, il grasso orbitale, o corpo adiposo dell'orbita, fornisce protezione ed isolamento all'occhio.
La tonaca fibrosa ha diverse funzioni: svolge la funzione di supporto meccanico e di protezione, fornisce l' inserzione sulla sclera per i muscoli estrinsechi dell'occhio. (e contiene strutture che coadiuvano la messa a fuoco degli oggetti)
La maggior parte di tutta la superficie oculare è ricoperta dalla sclerotica ( o sclera) , comunemente denominata “bianco dell'occhio”, formata da tessuto connettivo fibroso denso ed opaco, ricco di fibre collagene e di fibre elastiche, più spesso a livello del polo posteriore dell'occhio, in prossimità dell'uscita de nervo ottico, e più sottile nella sua porzione
anteriore.Le fibre hanno un decorso circolare e la loro particolare disposizione è fondamentale per l'elasticità e l'estensibilità.
La superficie della sclera contiene inoltre piccoli vasi sanguigni e nervi che raggiungono gli strati sottostanti.
La cornea è un tessuto trasparente e sottile, in continuità con la sclera e rappresenta la prima di un sistema di lenti (o mezzi diottrici) che viene percorsa dal raggio luminoso prima che venga formata l'immagine sulla retina; il confine con la sclera forma un bordo denominato limbus corneale. Il suo spessore, generalmente di 0,5mm circa,non è uguale per tutte e due gli occhi e tende a diminuire lievemente con l'età.
Ricopre un duplice compito, di protezione dell'occhio e di rifrazione dell'immagine, in quanto funziona come una vera e propria lente, come in una antica macchina fotografica.
La cornea non è vascolarizzata e la sue cellule epiteliali superficiali ricevono nutrimento e ossigeno dalle lacrime che scorrono sulla sua superficie libera. Al contrario, è riccamente innervata da terminazioni nervose libere ed è per questo che è la parte più sensibile dell'occhio.
La Tunica vascolare, o uvea, si compone della coroide, dell'iride e del corpo ciliare, e contiene molti vasi sanguigni e linfatici ed i muscoli instrinseci dell'occhio.
La coroide è lo strato riccamente vascolarizzato che separa la tunica fibrosa da quella nervosa.
Tale strato, ricoperto esternamente dalla sclera e unito internamente allo strato più esterno della retina, contiene una fitta rete capillare che trasporta ossigeno e nutrienti alla retina stessa.
E’ costituita inoltre da fibre connettivali e da cellule ripiene di un pigmento nerastro, i melanociti, che contribuiscono a mantenere la necessaria oscurità nella “camera oscura” endobulbare. Ed è proprio per il suo colore simile alla buccia dell’uva nera che viene anche denominata uvea.
L'iride, formata da materiale opaco, posto tra la cornea e il cristallino. Può essere osservata attraverso la superficie trasparente della cornea, e contiene vasi sanguigni, cellule pigmentate e due strati di fibre muscolari lisce che formano i muscoli pupillari. Sono proprio questi muscoli che quando si contraggono modificano il diametro della pupilla, cioè l'apertura centrale dell'iride, che in condizioni ordinarie appare di colore nero, in quanto la radiazione che raggiunge il fondo dell'occhio non ritorna indietro e non viene captata quindi dall'occhio di chi osserva.
Si potrebbe dire che l'iride si comporta come il diaframma di una macchina fotografica. La pupilla, paragonabile all'apertura di una macchina fotografica, regola la quantità di fasci di raggi luminosi, che, passando attraverso essa, vanno a colpire il fondo dell'occhio, cioè la retina.
Un gruppo di fibre muscolari lisce, disposte a formare cerchi concentrici intorno alla pupilla, quando entra troppa luce nell'occhio, si contraggono: è il cosiddetto muscolo sfintere (o costrittore) dell'iride (o della pupilla), che spinge l’iride verso il centro, provocando il restringimento del diametro della pupilla ( miosi ) e permettendo che una minore quantità di luce raggiunga la retina. Un secondo gruppo di fibre muscolari lisce, con orientamento radiale a partire dal margine pupillare, costituisce il muscolo dilatatore della pupilla. Quando la quantità di luce che entra nell'occhio diminuisce (come al buio o di notte), questo muscolo spinge l’iride dal centro verso l’esterno, causando la dilatazione della pupilla ( midriasi ) e permettendo così che una maggior quantità di luce raggiunga la retina. Il diametro della pupilla può variare generalmente tra i 2mm, in caso di forte illuminazione, e gli 8 mm, in caso di bassa illuminazione.
Entrambe questi muscoli sono sotto il controllo del sistema nervoso autonomo Per esempio, l'attivazione del sistema parasimpatico in risposta a una luce forte causa costrizione della pupilla (riflesso consensuale alla luce), mentre l'attivazione del sistema simpatico in risposta a una luce fioca determina la dilatazione pupillare.
Nella sua porzione periferica, l'iride si inserisce sulla parte anteriore del corpo ciliare, una ragione ispessita che si trova in profondità nel punto di passaggio fra sclera e cornea.
Posteriormente è in rapporto con la tunica nervosa, cioè con la retina, fino al punto dell'ora serrata, il punto di giunzione del corpo ciliare con la coroide.
Il corpo ciliare è formato dal muscolo ciliare e da numerose estroflessioni digitiformi dell'epitelio chiamate processi ciliari, dove prendono inserzione i legamenti sospensori del cristallino.
Quest'ultimo è posto dietro alla pupilla in modo che la luce quando penetra dalla pupilla, prima di giungere ai fotorecettori, attraversa anche il cristallino.
La tunica nervosa, o retina, è lo strato più interno dell'occhio. E' formata da due foglietti, uno esterno, chiamato parte pigmentata, a contatto con la coroide, e uno interno più spesso, chiamato parte nervosa, a contatto con il corpo vitreo, che è la parte fotosensibile dell'occhio. La parte pigmentata della retina assorbe la luce che ha attraversato il cristallino.
La parte nervosa, denominata anche retina propriamente detta, si distingue, dall'esterno verso l'interno, cioè verso il corpo vitreo, in altrettanti nove strati:
La parte nervosa è formata da molto recettori, cellule di supporto e cellule nervose che hanno il compito di elaborare ed integrare le informazioni visive.
I due strati della retina normalmente sono molto vicini fra loro, ma non sono colelgati strettamente. Infatti la parte pigmentata della retina prosegue anche sul corpo ciliare e sull'iride, mentre quella nervosa si interrompe in corrispondenza dell'ora serrata. La parte nervosa della retina ha dunque la forma di una coppa che avvolge l'occhio nella cavità posteriore.
Lo strato più esterno della retina, vicino alla parte pigmentata, contiene i fotorecettori, cioè le cellule in grado di percepire gli stimoli luminosi e trasformarli, attraverso reazioni biochimiche, in segnali elettrici, che istradati sulle vie ottiche, raggiungono le aree della visione della corteccia encefalica. L'occhio possiede due tipi di fotorecettori: coni e bastoncelli.
Nell'uomo, i coni e i bastoncelli non sono distribuiti in modo uniforme sulla supericie esterna della retina: il numero di bastoncelli è di gran lunga superiore a quello dei coni. Si ritiene che nella nostra retina ci siano circa 125 milioni di bastoncelli , localizzati principalmente in corrispondenza della periferia retinica (zona dell'ora serrata); spostandosi dalla periferia al centro della retina la densità dei bastoncelli diminuisce. I coni, circa 6 milioni, sono concentrati in un'area retinica corrispondente al punto in cui un'immagine visiva, dopo essere passata attraverso la cornea ed il cristallino, giunge a fuoco. Questa regione è chiamata anche macula lutea, e più dettagliatamente, la zona dove troviamo esclusivament coni è un'area chiamata fovea, o fovea centralis. La fovea è la zona della visione distinta, cioè quando fissimo un oggetto la sua immagine cade esattamente in tale zona. La linea teorica tesa fra l'oggetto e la fovea che passi esattamente per il centro del cristallino viene chiamata asse visivo dell'occhio.
La funzione dei bastoncelli è nettamente diversa da quella dei coni e preciasmente i primi non essendo in grado di discriminare i colori, sono così responsabili della visione notturna e crepuscolare. I coni per contro sono resposabili della percezione dei colori e precisamente sono deputati alla visione diurna, cioè della luce intensa e piena di colori. Esistono infatti tre tipi di coni, corrispondenti ai colori rosso, verde e blu, e la loro stimolazione nelle varie combinazioni consente la percezioni dei diversi colori. I coni constentono quindi una visione più nitida e distinta rispetto a quella offerta dai bastoncelli, necessitando però di una quantità di luce superiore. Ciò sarebbe in relazione alla presenza al loro interno di composti organici particolari, chiamati pigmenti visivi. L'assorbimento dei fotoni da parte di questi pigmenti è il passaggio chiave nella catena di eventi che costituisce il processo di fotoricezione, cioè di ricezione della luce.
I coni e i bastoncelli stabiliscono sinapsi con circa 6 milioni di neuroni, che vengono chiamate cellule bipolari, i quali a loro volta contraggono sinapsi con altri neuroni, le cellule gangliari.
L'organizzazione anatomica e funzionale della retina è di trasmettere l'impulso dai fotorecettori alle cellule bipolari grazie a gruppi di cellule orizzontali che si estendono nei punti di sinapsi. Nello strato in cui le cellule bipolari fanno sinapsi con le cellule gangliari si trovano le cellule amacrine. Quest'ultime insieme alle cellule orizzontali, hanno la funzione d facilitare o inibire la comunicazione fra i fotorecettori e le cellule gangliari e quella di modificare la sensibilità della retina.
Gli assoni di circa 1 milione di cellule gangliari convergono in una regione circolare, locallizzata medialmente rispetto alla fovea, che pende il nome di disco ottico o papilla (cfr paragrafo 2.3) e costituiscono il nervo ottico (N II), che le fibre attravers ano per portarsi verso il diencefalo. Questo è privo di tutti i fotorecettori e di tutte le strutture che caratterizzano la restante parte della retina e quando la luce colpisce questo punto non è percepibile e si parla di macchia cieca della retina, perchè compare nel campo visivo come una macchia bianca.
Come sopra citato, il corpo ciliare e il cristallino dividono l'interno dell'occhio in una cavità posteriore, più grande, chiamata anche camera vitrea, e in una cavità anteriore, più piccola. La cavità anteriore è, a sua volta, suddivisa in una camera anteriore che si estende dalla cornea all'iride, e in una camera posteriore, estesa dall'iride e dal corpo ciliare al cristallino. Le camere anteriore è piena di liquido, l'umor acqueo. La cavità posteriore contiene invece una sostenza gelatinosa nota come corpo vitreo o umor vitreo.
L'umor acqueo è un liquido che circola all'interno della cavità anteriore, passando dalla camera posteriore a quella anteriore attraverso la pupilla. Tale fluido diffonde liberamente attraverso il corpo vitreo e lungo la superficie retinica. Esso viene prodotto e secreto dall cellule epiteliali che rivestono i processi ciliari del corpo ciliare. Esso circolando all'interno delle camere dell'occhio, costituisce un'importante via per l'apporto di nutrienti e per lo smaltimento dei cataboliti e inoltre funge da ammortizzatore. Grazie alla pressione esercitata da questo liquido, l 'occhio mantiene la propria forma; infatti il liquido stabilizza la posizione della retina, in quanto comprime e fa aderire la parte nervosa a quella pigmentata.
L'umor acqueo viene secreto nella camera posteriore e riassorbito nella camera anteriore dove, dopo esser stato filtrato da una serie di ffibre collagene localizzate vicino alla base dell'iride, penetra all'interno del canale di Schlemm, o seno venoso della sclera, una sorta di canale che decorre circolarmente intorno all'occhio all'altezza del limbus.
La cavità posteriore dell'occhio contiene il corpo vitreo ( o umor vitreo), una massa gelatinosa che da una parte aiuta a mantenere la forma dell'occhio e dall'altra si deforma in risposta all'azione dei muscoli extraoculari nell'orbita. E' composto da cellule specializzate che producono le fibre collagene e i proteoglicani responsabili della consistenza gelatinosa di tale massa. Al contrario dell'umor acqueo, il corpo vitreo si forma durante lo sviluppo embrionale e non viene più rimpiazzato.
Nel sistema di mezzi diottrici sopra citato, rientra il cristallino (o lente) che si trova posteriormente alla cornea e viene mantenuto in sede grazie ai legamenti sospensori che originano dal corpo ciliare della coroide. E' costituito da una sostanza trasparente e molto elastica ed è composto da tre strati concentrici, di cui i due esterni, che sono, dall’esterno all’interno, rispettivamente la capsula e la zona corticale, circondano a mo’ di guscio, la parte più interna, il nucleo. In generale ha la forma di una lente biconvessa, di cui le curvature variano in base alla distanza dell'oggetto osseervato: questo processo è noto come accomodazione del cristallino, con il fine di focalizzare le immagini sulla retina.
Lungo il cristallino si trovano delle cellule denominate fibre del cristallino; si tratta di cellule che hanno perso il nucleo ed altri organuli, di forma sottile e allungata, piene di proteine trasparenti chiamate cristalline che sono responsabili sia della trasparenza che del potere focallizante del cristallino. Quindi la trasparenza del cristallino dipende dall'esatta combinazione e da caratteristiche biochimiche e strutturali. Quando tale equilibrio si altera il cristallino perde la propria trasparenza e tale condizione è nota come cataratta.
Gli annessi dell'occhio includono le palpebre, l'epitelio superficiale dell'occhio e le strutture adibite a produzione, secrezione e rimozione delle lacrime.
Le palpebre sono una continuazione della cute ed agiscono come un tergicristallo: grazie al loro continuo ammiccamento la superficie oculare viene lubrificata e vengono rimossi polvere e detriti. Esse possono anche rimanere serrate in modo da proteggere la superficie oculare. La rima palpebrale è lo spazio che separa i margini liberi della palpebra superiore e di quella inferiore.
Le ciglia sono dei peli piuttosto robusti che si inseriscono lungo il margine libero delle palpebre e contribuiscono a prevenire la deposizione di materiale estraneo ( inclusi gli insetti ) sulla superficie oculare.
Le ciglia sono associate a grandi ghiandole sebacee. Lungo il margine interno della palpebra, alcune ghiandole sebacee modificate, le ghiandole tarsali, o ghiandole del meibomio, secernono un prodotto ricco di liquidi che preveniene l'incollamento fra le due palpebre.
La cute che ricopre la superficie visibile delle palpebre è molto sottile e contiene in profondità fibre muscolari del muscolo orbicolare dell'occhio e del muscolo elevatore della palpebra superiore. Questi muscoli scheletrici sono responsabili, rispettivamente, della chiusura e dell'apertura della palpebra superiore.
L'epitelio che riveste la superficie interna delle palpebre e la superficie esterna del globo oculare viene chiamato congiuntiva, una membrana mucosa ricoperta da un particolare epitelio di tipo pavimentoso pluristratificato.
La cornea è ricoperta da un delicatissimo epitelio di tipo squamoso l'epitelio corneale, spesso da cinque a sette strati, in continuità con la congiuntiva oculare (che circonda il globo oculare).
Un costante apporto lacrimale bagna continuamente la superficie oculare, mantenendo la congiuntiva umida e detersa. Le lacrime riducono l'attrito e la dissidratazione, rimuovono il pulviscolo, prevengono le infezioni batteriche e forniscono nutrienti ed ossigeno all'epitelio congiuntivale.
Questo è consentito dall'apparato lacrimale che produce, distribuisce e rimuove le lacrime. E' costituito da una ghiandola lacrimale, con i dotti associati, canalicoli lacrimali, un sacco lacrimale e un dotto naso-lacrimale.
La ghiandola lacrimale normalmente produce gli ingredienti chiave e gran parte della quantità di lacrime necessario ad aumentare la superficie congiuntivale. I nutrimeti e l'oggiseno che sono necessari alle cellule corneali vengono normalmente forniti per diffusione dal film lacrimale (o liquido lacrimale) , acquoso e di PH alcalino.
Le lacrime contengono anche il lisozima, ossia un enzima che attacca i batteri, e diversi anticopri che attaccano i patogeni prima che penetrino nell'occhio.
La ghiandola lacrimale produce circa 1ml di lacrime ogni giorno. Una volta raggiunta la superficie oculare, le lacrime si mescolano al secreto delle ghiandole accessorie e a quello oleoso delle ghiandole tarsali, ne risulta un film acqua-olio che lubrifica e rallenta l'evaporazione. L'ammiccamento spande le lacrime attraverso la superificie oculare, accumulandole nell'angolo palpebrale mediale in un'area nota come lago lacrimale. Il lago lacrimale copre la caruncola lacrimale, che sporge anteriormente rispetto alla superficie oculare. I punti lacrimali sono due piccoli poli che drenano il lago lacrimale; essi riversano il contenuto all'interno dei canalicoli lacrimali, piccoli canali che convogliano le lacrimale nel sacco lacrimale, che alloggia nel solco lacrimale dell'orbita. Dalla porzione inferiore del sacco lacrimale si diparte il dotto naso-lacrimale, che si continua nel canale lasolacrimale, formato dalla sutura fra l'osso lacrimale e l'osso mascellare.
Il dotto naso-lacrimale trasporta le lacrime nella cavità nasale omolaterale e sbocca nel meato nasale inferiore, una stretta fessura in corrispondenza della porzione infero-laterale del cornetto nasale inferiore.
Le vie ottiche assicurano la trasmissione delle informazioni visive dall'occhio al cervello. Consistono in un insieme di strutture che, partendo dalla retina collegano il bulbo oculare alla porzione occipitale del cervello.
Rappresentano strutture sensoriali e percettive che permettono la visione delle immagini grazie alla ricezione di un impulso luminoso, il suo trasferimento sotto forma di impulso elettrico e la conseguente percezione ed interpretazione visiva.
Queste strutture sono rappresentate dal nervo ottico, il chiasma ottico, il tratto ottico, il corpo genicolato laterale e le radiazioni ottiche di Gratiolet.
Il nervo ottico è il II paio di nervi cranici, che origina dalla confluenza delle fibre ottiche provenienti dalla retina, in corrispondenza della papilla ottica (considerata la “testa” del nervo ottico). Le sue fibre sono ricoperte dalla mielina, e per tutto il suo decorso risulta avvolto dalle meningi: è per questo che viene considerato una continuazione del sistema nervoso centrale, come il nervo olfattivo (I).
Durante il suo percorso è possibile riconoscere quattro porzioni: quella intraoculare (papilla ottica), intraorbitario, intracanicolare (nel canale ottico) e il segmento intracranico.
Il nervo ottico, formato da fibre ottiche provenienti dalle cellule gangliari della retina, infatti, lascia l'orbita oculare passando per il canale ottico, scavato all'interno dello sfenoide; qui le sue fibre decussano e così il nervo ottico si dirige in direzione dell'emisfero controlaterale rispetto all'occhio di origine. Qui le fibre di entrambe i nervi ottici, provenienti dalla porzione mediale (o nasale) della retina e veicolanti le immagini provenienti dalla metà laterale del campo visivo, si incrociano con quelle del lato opposto, e formano il cosiddetto chiasma ottico, posizionato nella parte inferiore del cervello, immediatamente sotto al l'ipotalamo. Esistono tre tipi di fibre ottiche: le nasali, sopra citate, che decusanno interamente, le maculari, di cui metà decussano nel chiasma e l'altra metà prosegue diretta, e le temporali, che non si incrociano e continuano dirette.
Queste fibre, tutte insieme, fuoriescono posteriormente al chiasma e formano i tratti ottici che contraggono sinapsi all'interno del corpo genicolato laterale del talamo, dove terminano la maggior parte delle fibre. Da qui originano fibre talamo- corticali, che formano le radiazioni ottiche di Gratioleet, dirette alla corteccia visiva primaria.
La porzione di fibre che terminano in prossimità del talamo si portano direttamente nei collicoli superiori del mesencefalo.
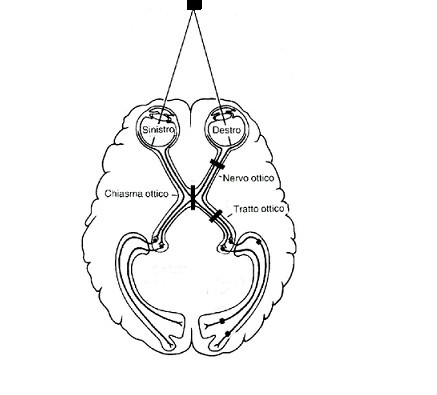
Fig. 2 - Le vie ottiche
Le fibre nervose delle radiazioni ottiche di Gratiolet decorrono nella sostanza bianca, raggiungono la scissura calcarina e terminano nella sostanza grigia dell’area visiva (V 1) , nel lobo occipitale. L’area corticale relativa (denominata Area striata o Area 17 di Brodmann) si estende fino in corrispondenza della porzione posteriore del corpo calloso e rappresenta il recettore centrale sede delle sensazioni visive elementari. E' deputata alla percezione cosciente degli stimoli visivi, cioè all'interpretazione dei segnali elettrici prodotti dalla stimolazione dei fotorecettori sulla retina.
La corteccia visiva primaria di ogni emifero riceve le vie sensitive provenienti dall'occhio controlaterale e quindi i segnali riguardanti il campo visivo dell'emilato opposto, e a sua volta invia comandi motori alla porzione del corpo controlaterale.
Ogni regione encefalica, sia essa motoria o sensitiva, è connessa a delle aree dette associative, ovvero delle regioni corticali che monitorano ed interpretanogli impulsi rispettivamente motori o sensitivi che giungono alla corteccia primaria. Tra queste ritroviamo le aree associative visive.
L'aree associative visive controllano gli impulsi della corteccia visiva primaria e ne interpretano e ne integrano i risultati. In sostanza, con un esempio pratico, ci consentono di riconoscere le lettere dell'alfabeto e la parola di senso compiuto formata dalla successione di più lettere. Una lesione a queste aree, ci permetterebbe di scansionare le linee o righe di lettere in una pagina stampata, ma non saremmo in grado di attribuirle un significato.
Precisamente si tratta di aree extrastriate: l'area parastriata o area 18 di Brodmann ( V2 ) , che a sua volta si continua esternamente con l’area peristriata o area 19 di Brodmann ( V3 ).
Altre due aree con funzioni simili, che ritroviamo nella figura (numero della figura da decidere) sono: l'area V 4, deputata al riconoscimento di colori e forme e l'area V 5, conosciuta come corteccia infero-temporale, una regione della corteccia visiva extrastriata deputata alla percezione del movimento e nella guida di alcuni movimenti oculari.
Dalla corteccia primaria si dipartono due vie di propagazione dell'impulso visivo attraverso il cervello: la via ventrale e la via dorsale. La prima, detta anche “via del cosa” è deputata al riconoscimento degli oggetti e delle forme e nell'immagazzinamento nella memoria a lungo termine; durante il suo percorso attraversa le aree V 2 e V 4e termina nel lobo temporale inferiore.
La via dorsale, detta anche “via del dove”, percorre l'area V 2 e V 5 e giunge fino alla corteccia parietale posteriore; è associata al movimento e alla rappresentazione degli oggetti nello spazio e al controllo occhio-mano mentre si tenta di afferrare un oggetto.
Oltre al nervo ottico, citato nei paragrafici precedenti, altre paia di nervi cranici sono deputati a svolgere funzioni inerenti all'occhio. Tra questi ritroviamo il nervo oculomotore, il nervo trocleare, una parte del nervo trigemino, ed il nervo abducente.
Il nervo oculomotore ( il III paio di nervi cranici) è un nervo deputato ai movimenti oculari; ha origine nel mesencefalo e penetra nella parte posteriore dell'orbita. E' destinato all'innervazione dei muscoli estrinsechi dell'occhio, che servono per i movimenti del bulbo oculare, e al muscolo elevatore della palpebra superiore. I pazienti affetti da lesioni di questo nervo riportano ptosi palpebrale e vedono doppio in quanto, non essendo i movimenti dei due occhi appropriatamente coordinati, si creano immagini retiniche non corrispondenti.
Il nervo trocleare (IV) è un nervo motore, che origina dal mesencefalo e innerva il muscolo obliquo superiore di ciascun occhio. Prende il nome dalla troclea, un legamento del muscolo prima citato. Lesioni a questo nervo comportano difficoltà a guardare verso il basso e lateralmente.
Il nervo trigemino ( V ) è il nervo misto, sensitivo e motorio, della faccia. Una sua porzione origina dalla branca oftalmica e ne innerva le strutture, ossia i componenti dell'orbita, la cornea, le cavità nasali, la plapebra superiore, le sopracciglia e la cute della fronte.
Infine il nervo abducente (VI) è anch'esso un nervo motore, che origina dal ponte encefalico e raggiunge l'occhio attraverso la fessura orbitaria superiore omolaterale, come i nervi III e IV. E' deputato all'innervazione dei muscoli retti laterali dell'occhio.
Il sistema oculomotore controlla i movimenti degli occhi all'interno delle orbite. I movimenti oculari hanno un duplice scopo: mantenere sulla fovea l'immagine costante di oggetti in movimento e dirigere la fovea sui diversi particolari dell'ambiente per costruire un immagine completa della realtà.
Essendo l'occhio una sfera capace di ruotare su se stessa, possiamo riconoscere tre tipologie di movimento, intorno ai tre assi: movimenti orizzontali, movimenti verticali e movimentitorsionali.
Inoltre, se gli assi visivi durante il movimento vengono mantenuti nella stessa direzione, si parlerà di movimenti coniugati (o versioni); qualora i movimenti oculari avvengano invece in direzioni opposte, si parlerà di disgiunzioni (o vergenze).
Infine, in base alla velocità del movimento, possiamo distinguere movimenti lenti o rapidi: si tratta in entrambe i casi di movimenti attuati dai nuclei dei nervi motori dello'occhio.
Fra i movimenti rapidi ritroviamo il movimento saccadico che porta velocemente, come il movimento della fase rapida del nistagmo, la fovea sull'oggetto di interesse.
Si tratta di movimenti programmati, stereotipati e che possono rispondere a stimoli tattili causati volontariamente o anche a ricordi inconsci della posizione di un oggetto nello spazio. Il movimento saccadico, è in grado di adattarsi alla variazione dello stato funzionale dei muscoli estrinsechi degli occhi: infatti, per esempio, nel caso di paresi di un muscolo, il movimento saccadico diventa ipometrico, e se si occlude un occhio, automaticamente il saccadico dell'occhio sano diventa ipermetrico.
Il controllo di questa attività avviene a livello corticale (campi oculari frontali, nucleo caudato), più precisamente nel collicolo superiore dove vengono integrate le informazioni visive provenienti dalle vie ottiche, e nel cervelletto: per esempio lesioni del verme cerebellare interferisconono con l'ampiezza dell'impulso e con l'adeguamento dell'intensità degli step saccadici all'intensità dell'impulso.
Fra i movimenti lenti ritroviamo la fase lenta del nistagmo e l'inseguimento lento. Questi provocano il movimento degli occhi nello spazio per mantenere la fovea su un bersaglio visivo, calcolano la velocità del movimento e fanno muovere entrambe gli occhi alla stessa velocità, in modo da mantenere una visione perfetta. Alcuni studi mostrano l'attivazione di alcune cellule della corteccia parietale durante questi movimenti.
I “semplici” movimenti oculari sottendono in realtà ad un sistema neuroanatomico molto complesso che garantisce e mantiene una perfetta visione binoculare: il sistema oculomotore. I neuroni oculomotori sono differenti dai motoneuroni spinali, in quanto tutti i neuroni prendono parte a tutti i tipi di movimento oculari e non sono specializzati nel singolo movimento, non rispondono a variazioni impreviste a carico dell'ambiente esterno e hanno modalità di risposta stereotipate.
Il sistema oculomotore è strettamente in connessione con il sistema visivo e con quello cognitivo, ed è grazie a questa integrazione che ci è permesso individuare gli oggetti e localizzarli nello spazio, identificare i movimenti e percepire le profondità: la stessa visione binoculare è il risultato della cooperazione fra questi sistemi.
Il sistema oculomotore riconosce diversi tipi di movimento, sotto elencati.

Fig.3 - Muscolatura estrinseca dell'occhio
I muscoli oculari estrinsechi, o muscoli oculomotori, che provvedono ai movimenti del bulbo oculare, alcuni citati nel paragrafo precedente, sono in numero sei: i quattro retti, mediale, laterale, superiore ed inferiore e i due obliqui superiore ed inferiore, e ad essi viene solitamente aggiunto il muscolo elevatore palpebrale.
Le fibre muscolari che ricoprono circa la totalità della lunghezza di questi muscoli, sono di due tipi: sottili a risposta lenta, che occupano la parte periferica del muscolo e spesse a risposta rapida, situate nella porzione interna.
Tutti i muscoli possiedono due inserzioni, una bulbare e una orbitaria, che si realizzano tramite strutture tendinee di diversa lunghezza.
I tendini prossimali dei quattro muscoli retti prendono origine precisamente dalla porzione posteriore dell'orbita, da un ispessimento periosteo chaimato anello di Zin, che circonda il canale ottico e la parte centrale della fessura orbitaria superiore. Hanno inserzioni rispettivamente sulla superficie mediale, laterale, superiore ed inferiore del bulbo oculare, nella sclera. I tendini di questi muscoli contengono organi di Golgi modificati, estreaamente sensibili alla tensione muscolare. Questo, insieme ai fusi neuromuscolari contenuti nel corpo del muscolo e sensibili alle sue variazioni di lunghezza, inviano al tronco dell'encefalo informazioni fondamentali per il controllo dell'oculomotricità.
Il tendine prossimale dell'obliquo superiore origina dal periostio dello sfenoide in prossimita dell'anello di Zin e al lato del muscolo elevatore; quello dell'obliquo inferiore origina invece dall'osso mascellare a livello della parte anteriore dell'orbita e lateralmente al canale naso lacrimale.
|
Muscolo |
Origine |
Inserzione |
Azione |
Innervazione |
|
Retto inferiore |
Sfenoide (intorno al canale ottico) |
Superficie inferiore del bulbo oculare |
Guardare in basso |
Nervo oculomotore |
|
Retto mediale |
Sfenoide (intorno al canale ottico) |
Superficie mediale del bulbo oculare |
Ruotare medialmente l'occhio |
Nervo oculomotore |
|
Retto superiore |
Sfenoide (intorno al canale ottico) |
Superricie superiore del bulbo oculare |
Guardare in alto |
Nervo oculomotore |
|
Retto laterale |
Sfenoide (intorno al canale ottico) |
Superficie laterale del bulbo oculare |
Ruotare lateralmente l'occhio |
Nervo abdcente |
|
Obliquo inferiore |
Osso mascellare, a livello della porzione anteriore dell'orbita |
Superficie laterale inferiore del bulbo oculare |
Rotazione oculare, guardare in alto e lateralmente |
Nervo oculomotore |
|
Obliquo superiore |
Sfenoide (intorno al canale ottico) |
Superifice laterale superioer del bulbo oculare |
Rotazione oculare, guardare in basso e lateralmente |
Nervo trocleare |
Tab. 2 I muscoli estrinsechi dell'occhio.























